
ავტორი: გელა თევზაძე, ბიოლოგიურ მეცნიერებათა დოქტორი, თავისუფალი უნივერსიტეტის პროფესორი
ჩვენს ბლოგზე განთავსებულმა თემამ “შეიძლება თუ არა მართლმადიდებელი იყოს ევოლუციონისტი?” ცხარე კამათი გამოიწვია, სადაც რამდენიმე კითხვა იქნა დასმული ევოლუციის ოპონენტთა მხრიდან. ვითვალისწინებთ რა ჩვენი მკითხველის თვალსაზრისს, გვსურს დავაკმაყოფილოთ მათი სურვილი და ამჯერად გთავაზობთ თავისუფალი უნივერსიტეტის პროფესორის, ბიოლოგიურ მეცნიერებათა დოქტორის გელა თევზაძის სამ სტატიას, რომელიც გამოქვეყნდა გაზეთ 24 საათის სამეცნიერო პოპულარულ სტატიათა რუბრიკაში სამ ნაწილად. გამომდინარე იმ კითხვებიდან რაც ჩვენი ბლოგის ფეისბუკის ფორუმზე დაფიქსირდა, აღნიშნული თემა წმინდა მეცნიერულ-პოპულარული ხასიათისაა და ავტორი საკითხს მხოლოდ ამ კონტექსტში განიხილავს. თითოეულ სტატიას რამდენიმე ილუსტრაციაც ახლავს, რომელთა მოკლე აღწერა ყოველი სტატიის ბოლოშია წარმოდგენილი. აღნიშნული სტატიები საინტერესოა იმდენად, რამდენადაც, თუ დიაკონ ანდრეი კურაევის მონოგრაფია “შეიძლება თუ არა მართლმადიდებელი იყოს ევოლუციონისტი?” საკითხს საღვთისმეტყველო თვალსაზრისით განიხილავს, წინამდებარე სტატიები პრობლემას მეცნიერული კუთხით აშუქებენ, რაც ერთგვარად ავსებს ხსენებული მონოგრაფიის შინაარსს.
სტატია ქვეყნდება ავტორის ნებართვით, რისთვისაც მადლობას ვუხდით მას ცენტრის სახელით
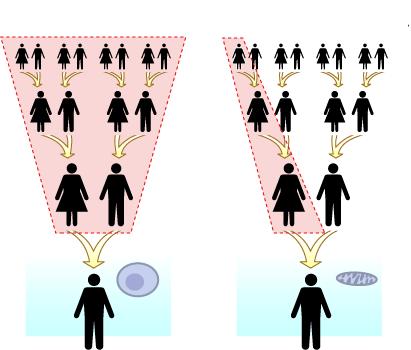
მიტოქონდრიული დნმ:
არაქრომოსომული გენეტიკური ინფორმაცია,
რომელიც ჩვენი წარმომავლობის ისტორიას შეიცავს.
ცნობილია,რომ ყოველი ცოცხალი ორგანიზმი უჯრედებისაგანშედგება. ყველა მრავალუჯრედიანი (და ზოგიერთი ერთუჯრედიანი) ორგანიზმის უჯრედებიშეიცავს ბირთვსა და მის გარშემოარსებულ ციტოპლაზმას. ორგანიზმების გენეტიკური ინფორმაციადეზოქსირიბონუკლეინის მჟავაში (დნ მჟავაში, ან დნმ–ში) არის კოდირებული, მისი უდიდესი ნაწილი უჯრედების ბირთვისშიგნით არის მოთავსებული და ქრომოსომებშია განლაგებული. მნიშვნელოვანიგამონაკლისია მიტოქონდრიული დნ მჟავა,რომელიც განლაგებულია ბირთვის გარეთ, ციტოპლაზმაში, უფრო ზუსტად კიმიტოქონდრიებში. მიტოქონდრიები ციტოპლაზმაში არსებული უჯრედსშიდა წარმონაქმნებია,რომელთა მთავარი ფუნქცია უჯრედისათვის აუცილებელი ენერგიის წარმოქმნა დადაგროვებაა.
რადგანმიტოქონდრიები – და შესაბამისად, მათში მოთავსებული დნმ – ბირთვის გარეთ არსებობს,მათი მემკვიდრეობით გადაცემის მექანიზმი მნიშვნელოვნად განსხვავდება ბირთვულიდნმ–ის გადაცემის მექანიზმისაგან. როგორც ცნობილია, ყველა ორგანიზმი ბირთვულიგენეტიკური მასალის ორ ტოლ ნახევარს იღებს: ერთს – დედისაგან, მეორეს კი –მამისაგან. ამიტომ, მაგალითად, ყველა ადამიანის 46 ქრომოსომიდან 23 დედისგანაამიღებული, 23 კი – მამისაგან. ეს მოვლენა იმაზეა დაფუძნებული, რომ სასქესო უჯრედისწარმოქმნისას მასში ორგანიზმის ბირთვული გენეტიკური მასალის (ბირთვული დნმ–ის)რაოდენობის ნახევარი გადადის. განაყოფიერებისას, ანუ მდედრობითი და მამრობითისასქესო უჯრედების შერწყმისას, ეს ორი ნახევარი (ადამიანის შემთხვევაში – 23–23ქრომოსომა) ერთიანდება და იწყება ახალი ორგანიზმის განვითარება.
მიტოქონდრიულიდნმ–ის შემთხვევაში მემკვიდრეობის მექანიზმი მნიშვნელოვნად განსხვავებულია. ესგანსხვავება იმითაა განპირობებული, რომ, თუმცა მდედრობითი და მამრობითი სასქესოუჯრედები ბირთვული დნმ–ის შემცველობის მხრივ ტოლფასნი არიან, ისინი მკვეთრადგანირჩევიან ციტოპლაზმის (და, მაშასადამე, ციტოპლაზმაში არსებული მიტოქონდრიულიდნმ–ის) შემცველობით: კვერცხუჯრედი (მდედრობითი სასქესო უჯრედი) ციტოპლაზმას დიდირაოდენობით შეიცავს, სპერმატოზოიდი (მამრობითი სასქესო უჯრედი) კი ციტოპლაზმასთითქმის მთლიანადაა მოკლებული. შესაბამისად, ყოველ ორგანიზმში არსებულიმიტოქონდრიული დნმ სწორედ დედის მხრიდანაა მიღებული.
ესგარემოება განსაზღვრავს მიტოქონდრიული (და ზოგადად, ციტოპლაზმური) დნმ–ისმემკვიდრეობით გადაცემის თავისებურებას: მიტოქონდრიულ დნმ–ში კოდირებულიინფორმაცია დედისაგან გადაეცემა როგორც ბიჭებს, ასევე გოგონებს, მაგრამ ამ ინფორმაციის გადაცემა შემდეგი თაობისათვისმხოლოდ ქალებს შეუძლიათ. გავრცელებული მეტაფორით, ყველა ადამიანი თავის უჯრედებში„ევას მიტოქონდრიულ დნმ–ს“ ატარებს. იგივე მეტაფორას თუ განვავრცობთ, ადამისმიტოქონდრიული დნმ პირველივე თაობაში „დაიკარგა“.
ექსპერიმენტულმეცნიერებაში კი ადამიანის ევოლუციის, მისი წარმომავლობისა და დედამიწაზეგანსახლების ქრონოლოგიის შესწავლისას, მიტოქონდრიული დნმ–ის ანალიზი უკვე მიღებულიდა აღიარებული მეთოდია. თუ ბირთვული დნმ ყოველ თაობაში განიცდის ცვლილებას(ქრომოსომების ფრაგმენტების მიმოცვლის, ახალი ორგანიზმის წარმოქმნისას ახლებურიკომბინაციების წარმოქმნის გამო), მიტოქონდრიული დნმ უცვლელად გადაეცემა შვილებსდედისაგან, და ქალიშვილებისაგან – შვილიშვილებს. მემკვიდრეობითი ცვლილებებიმიტოქონდრიულ დნმ–ში მეტად იშვიათია, რადგან, ბირთვული დნმ–ისაგან განსხვავებით,იგი არც ფრაგმენტებს მიმოიცვლის და არც ორად იყოფა სქესობრივი უჯრედებისწარმოქმნისას, რომ შემდეგ ახალ „ნახევარს“ შეერწყას. ის ცვლილებები კი, რომელიცმიტოქონდრიულ დნმ–ში მაინც ხდება, ძალიან ინფორმატულია და ადამიანთა დიდი ჯგუფებისწარმომავლობის ერთგვაროვნებისა თუ განსხვავებულობის მიმანიშნებელია.
მიტოქონდრიულიდნმ–ის გამოყენება ადამიანის წინაპრებისა და მისი წარმომავლობის შესასწავლად 1987წელს დაიწყო, როდესაც რებეკა კანმათანამშრომლებთან ერთად გამოაქვეყნდა სტატია „მიტოქონდრიულიდნმ და ადამიანის ევოლუცია“ (Rebecca Cann, “Mitochondrial DNA and HumanEvolution”). ამ სტატიაში გადმოცემული იყო 147 ადამიანის მიტოქონდრიულიდნმ–ის ანალიზი. ამ ადამიანების წარმოშობა (უფრო ზუსტად, მათი მდედრობითი სქესისწინაპრების წარმომავლობა) ხუთ სხვადასხვა გეოგრაფიულ რეგიონს მიეკუთვნებოდა.სტატიის ორი ძირითადი დებულება, რამდენიმე მცირე უზუსტობის მიუხედავად, ახლაცჭეშმარიტადაა მიჩნეული: 1) „მიტოქონდრიული დნმ–ის ხეს“ ფესვები აფრიკაში აქვსგადგმული, ანუ ხუთივე რეგიონის წარმომადგენლების წარმომავლობა აფრიკაში იწყებოდა;2) ამ ხის ყველა „ტოტი“ შედარებით მოკლეა, რაც მიგვანიშნებს, რომ ხუთივე ჯგუფისსაერთო წინაპარი შედარებით მცირე ხნის წინ ცხოვრობდა. ცხადია, რომ ამ საერთოწინაპრის დასახელებისათვის „ევა“ ყველაზე შესაფერი აღმოჩნდა…
ამდებულებების საფუძველზე, ავტორებმა დაასკვნეს, რომ მათი შედეგები კარგად მიესადაგებოდაადამიანის ევოლუციის ერთ–ერთ მოდელს. ამ მოდელის თანახმად, ანატომიურადთანამედროვე ადამიანის წარმოშობა პირველად აფრიკაში მოხდა თითქმის 150 ათასი წლისწინათ; ამას მოჰყვა ადამიანების განსახლება აფრიკიდან – დაახლოებით 100 ათასი წლისწინ. ამ განსახლების შედეგად, აფრიკიდან გამოსულმა ადამიანებმა სრულიად შეცვალეს(განდევნეს) ყველა სხვა ჯგუფი, რის შედეგადაც ყველა თანამედროვე ადამიანი ამ –აფრიკაში წარმოშობილი და შემდგომ განსახლებული – პოპულაციის (ანუ ინდივიდთაჯგუფის) შთამომავალია. ეს ჰიპოთეზა, მიტოქონდრიული დნმ–ის კვლევის შედეგებისაგანდამოუკიდებლად, უკვე განიხილებოდა პალეოანთროპოლოგიურ ლიტერატურაში. მისი მთავარიალტერნატივა იყო ე.წ. მულტირეგიონალური(მულტიცენტრული) ჰიპოთეზა, რომლისთანახმადაც როგორც აფრიკაში, ასევე ევრაზიაში მოსახლე „წინაპართა პოპულაციები“მონაწილეობდნენ ანატომიურად თანამედროვე ადამიანის ფორმირებაში.
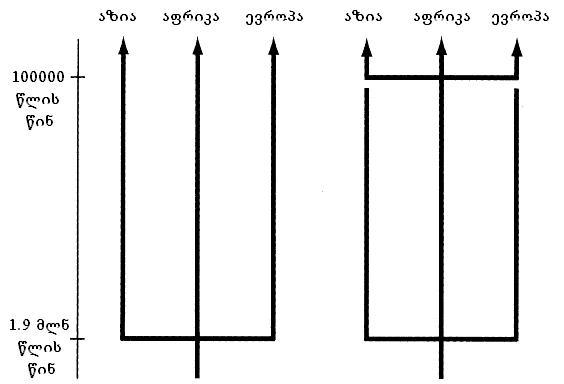
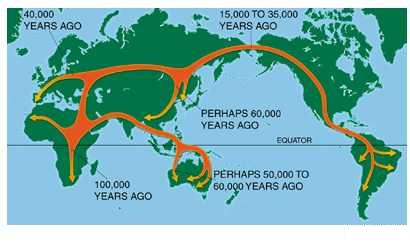
1989წელს უკვე გამოიცა პირველი სახელმძღვანელო ადამიანის ევოლუციაში, სადაც რებეკაკანის შედეგები იყო ნახსენები. სურათზე, რომელიც ამ სახელმძღვანელოდააგადმობეჭდილი, ორივე ხსენებული ჰიპოთეზა გრაფიკულადაა მოცემული. როგორცილუსტრაციიდან ჩანს, ორივე ჰიპოთეზის მიხედვით ადამიანის წარმოშობა აფრიკაში იღებსსათავეს, განსახლება კი 1.7–1.9 მილიონი წლის წინ დაიწყო. ადამიანის წინაპრებს, რომლებიც იმ დროშიცხოვრობდნენ, Homo erectus–ს(„გამართულ ადამიანს“) უწოდებენ. პირველი (მულტირეგიონალური) მოდელის თანახმად,აფრიკიდან Homo erectus –ის სამიგანსხვავებული პოპულაცია გამოვიდა და მათგან ადამიანის თანამედროვე აფრიკელები,აზიელები და ევროპელები ჩამოყალიბდნენ. მეორე მოდელის ვარაუდით, პირველი განსახლება მართლაც 1.7–1.9 მილიონიწლის წინ შეიძლება მომხდარიყო, მაგრამბევრად გვიან (დაახლოებით 100 ათასი წლის წინ) აფრიკიდან კიდევ ერთი ფორმაგამოვიდა, რომელმაც ევროპასა თუ აზიაში ყველა დანარჩენი პოპულაცია შეცვალა(გამოდევნა). ეს პროცესი მეორე მოდელის ამსახველ ნახაზზე (B) წყვეტილიმონაკვეთითაა აღნიშნული. ამ მოდელს, პირველისაგან ძირითადი განსხვავებისაღსანიშნავად, „აფრიკიდან ჩანაცვლების“(Out-of-Africa Replacement) მოდელს უწოდებენ.
რებეკაკანის ნაშრომში აღნიშნულია, რომ მათი შედეგების საფუძველზე აგებული „მიტოქონდრიულიხის“ „ტოტები“ არააფრიკულ (ევროპულ, აზიურ) პოპულაციებს შორის მკვეთრადგანსხვავებული წინაპრების არსებობას არ მიანიშნებდნენ. ასეთი წინაპრები კი,პირველი მოდელის თანახმად, ნამდვილად უნდა არსებობდნენ! ამგვარად, გენეტიკური კვლევისშედეგებმა უარჰყო მულტირეგიონალური ჰიპოთეზა, „აფრიკიდან ჩანაცვლების“ ჰიპოთეზას კი მხარს უჭერდა. სულ ცოტა ხანში,სასკოლო სახელმძღვანელოებისა და სამეცნიერო–პოპულარული ლიტერატურის თანახმად, „აფრიკიდანჩანაცვლება“ ადამიანის ევოლუციის ერთადერთ ჭეშმარიტ მოდელად იქნა მიჩნეული.
მაგრამაუცილებელია აღინიშნოს, რომ რებეკა კანის სტატიაში რამდენიმე უმართებულო მსჯელობაარსებობს: 1) მის მიერ აღწერილი პირველი მოდელი სრულყოფილად ვერ აკმაყოფილებს„მულტირეგიონალური“ ჰიპოთეზის განსაზღვრებას (დაწვრილებით იხ. ქვემოთ); 2) კანსადა მისი თანაავტორებს თავისი ჰიპოთეზა ექსპერიმენტულად (მაგ., სტატისტიკურიანალიზის გამოყენებით) არ შეუმოწმებიათ; 3) არსებობს ადამიანის ევოლუციის არა მხოლოდორი, არამედ რამდენიმე მოდელი, ასე რომ ორიდან ერთის უარყოფა სულაც არ ნიშნავს,რომ მეორე მოდელის ჭეშმარიტება ეჭვსგარეშეა. 4) ზოგადი პროცესის სტატისტიკურიდასაბუთება არ უზრუნველჰყოფს ამ პროცესის შემადგენელი ნაწილების ამსახველიმოდელების ჭეშმარიტების დამტკიცებას. ახლაეს ოთხი სუსტი წერტილი ცალკ–ცალკე განვიხილოთ:
1) რა არის მულტირეგიონალური ჰიპოთეზა? ადამიანის ევოლუციის მულტირეგიონალური მოდელიუშვებს, რომ შესაძლებელია ევოლუციის გასაოცარი პარალელიზმი ადამიანის წინაპრებიდანმის – სულ ცოტა, სამ – თანამედროვე ფორმებამდე. ეს მოდელი ჯერ კიდევ კანისნაშრომის გამოქვეყნებამდე იყო დისკრედიტირებული, თუმდაც იმის საფუძველზე, რომასეთი „სამგზის პარალელური ევოლუცია“ თეორიულად ძნელი წარმოსადგენი იყო. მეტიც,კვლევებმა ცხადჰყო, რომ ევროპელ, აზიელ და აფრიკელ პოპულაციებს შორის არსებულიგენეტიკური განსხვავებები მცირე იყო და არ იძლეოდა იმის საფუძველს, რომ ძველი,ერთმანეთისაგან მნიშვნელოვნად დაშორებული და დამოუკიდებელი წინაპრების არსებობადაგვეჯერებინა. ამიტომ, კანის სტატიაში განხილული მოდელი იმ დროისათვისაც კი უკვეუარყოფილი იყო. მაგრამ უკვე არსებობდა სხვაგვარი მულტირეგიონალური მოდელი, რომლისთანახმადაც ადამიანთა სხვადასხვა პოპულაციები (ავსტრალიური, მონღოლური,სამხრეთაფრიკული, ევრაზიული) ევოლუციის დროს აქტიურად მიმოიცვლიდნენ გენებს (ე.წ.„გენების დრეიფის“ მექანიზმით). სხვაგვარად რომ ვთქვათ, ხდებოდა არა „სამგზის“ (ან„ოთხგზის“!) პარალელური ევოლუცია, არამედ ურთიერთდაკავშირებული განვითარება.თითოეული პოპულაცია, რასაკვირველია, დანარჩენებისაგან მეტ–ნაკლებად განსხვავდება, მაგრამ ერთმანეთისაგანსრულიად დამოუკიდებელ ევოლუციას ეს მოდელი არც კი ახსენებს. მეტიც, ამ მოდელშიოთხი „ევოლუციური ხაზი“ მუდმივ ურთიერთქმედებაში იმყოფება და ცალკეულ განშტოებებსარ წარმოქმნის. და ბოლოს, „მულტირეგიონალური მოდელის“ არა ერთი, არამედ რამდენიმევერსია არსებობს, რომლებიც განსხვავდებიან ევოლუციის პროცესში გენების დრეიფისა დაგადარჩევის პირობების როლების სხვადასხვაგვარი განმარტებით. „აფრიკიდანგანსახლება–გამოცვლის“ ჰიპოთეზა კი „მულტირეგიონალური მოდელებთან“ შედარებით უფრომარტივად ხსნის ადამიანის ევოლუციაში მიმდინარე პროცესებს. სწორედ ამ მოსაზრებითაააღიარებული „აფრიკიდან ჩანაცვლების“ მოდელი ადამიანის ევოლუციის აღწერისათვისყველაზე უფრო შესაფერის საწყის ჰიპოთეზად.
2. ჰიპოთეზასთან თავსებადობა თუ ჰიპოთეზისშემოწმება? ყველა სტუდენტისათვის მეტადმნიშვნელოვანია იმის გაცნობიერება, რომ მეცნიერების ერთ–ერთი უმთავრესი მიზანიაექსპერიმენტული შედეგების დაგროვება რაიმე ჰიპოთეზის შესამოწმებლად. სამწუხაროდ,მრავალ შემთხვევაში ადამიანის წარმომავლობასთან დაკავშირებით ჩატარებულ გენეტიკურექსპერიმენტებში მკვლევარები თავს არიდებენ ჰიპოთეზის შემოწმებას და მხოლოდ იმასაჩვენებენ მკითხველს, რომ მიღებული შედეგები თავსებადია რომელიმე (ხშირად ადრევე„შერჩეულ“) ჰიპოთეზასთან. როგორც აღვნიშნეთ, საწყისი შედეგები მიტოქონდრიულიდნმ–ის გამოყენებით თავსებადი იყო როგორც „გამოცვლის“ მოდელთან, ასევე„მულტირეგიონალური მოდელის“ ახალ ვერსიასთან. ძირითადი სხვაობა ამ მოდელებს შორისიმაში მდგომარეობდა, მართლაც მოხდა თუ არა ევრაზიული პოპულაციების „ჩანაცვლება“.სხვა მხრივ – აფრიკული „ფესვებისა“ თუ შედარებით მცირე ხნის წინ არსებული საერთოწინაპრის თვალსაზრისით – ეს ორი მოდელი ერთმანეთისაგან თითქმის არ განსხვავდებოდა.ჩანაცვლების დასაბუთებისათვის კი აუცილებელია, რომ მიღებული შედეგები საკმარისინფორმაციას გვაძლევდნენ პოპულაციების შესახებ როგორც ჩანაცვლების პერიოდში და მისშემდეგ, ასევე იმ დრომდეც, როცანავარაუდევი ჩანაცვლება მოხდა. მხოლოდ ამ შემთხვევაში შესაძლებელია საუბარი იმჰიპოთეზის შემოწმებაზე, რომლის თანახმადაც რაიმე ჩანაცვლება მართლაც მოხდა.მაშასადამე, ჭეშმარიტად ფასეულ ინფორმაციას გენეტიკური კვლევის ის შედეგებიშეიცავენ, რომელთა საფუძველზე შესაძლოა ცვალებადობის ხარისხის დადგენა ჩანაცვლებამდე, ჩანაცვლებისას და ჩანაცვლებისშემდეგ, და ამ მონაცემების შედარება. კანის კვლევაში კი, სამწუხაროდ, ასეთიშედეგები არ იყო მიღებული…
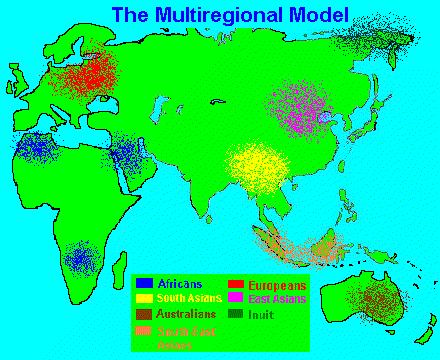
3. ბირთვული გენების „ხის ფესვები“. კანის ნაშრომში „ჩანაცვლების“ ჰიპოთეზის ერთ–ერთუმთავრეს მხარდამჭერ საბუთად მიტოქონდრიული დნმ–ის „ხის“ „აფრიკული ფესვები“ითვლება. მაგრამ „აფრიკული ფესვები“ ხომ მულტირეგიონალურ მოდელთანაცაა თავსებადი?თუ ამ უკანასკნელი მოდელის განხილვისას დავუშვებთ, რომ ადამიანთა წინაპრებისძირითადი პოპულაცია აფრიკაში ~2–1.5 მილიონი წლის წინაც ცხოვრობდა, „აფრიკულიფესვები“ აქაც სავსებით ბუნებრივი მოვლენაა…
რამდენიმეხნის შემდეგ ჩატარდა ანალოგიური კვლევა Y-ქრომოსომაზე. ეს ქრომოსომა მხოლოდმამაკაცებს აქვთ და, შესაბამისად, მამებისაგან მხოლოდ მამრობითი სქესის შვილებსგადაეცემათ. მიუხედავად იმისა, რომ Y-ქრომოსომა ბირთვში მდებარეობს, იგი, სხვაქრომოსომებისაგან განსხვავებით, გამრავლებისა და ახალი ორგანიზმების წარმოქმნისდროს არ მიმოიცვლის ფრაგმენტებს და თითქმის უცვლელი ხდება თაობათა განმავლობაში.სხვაგვარად რომ ვთქვათ და ამ სტატიის დასაწყისში გამოყენებული მეტაფორამოვიშველიოთ, ყველა მამაკაცს „ადამის Y-ქრომოსომა“ აქვს. ამიტომაც Y-ქრომოსომისანალიზი მიტოქონდრიული დნმ–ის ტოლფასადშეიძლება ჩაითვალოს.
Y-ქრომოსომისანალიზით მიღებული შედეგები ასევე თავსებადი იყო „აფრიკული ფესვების“ არსებობის იდეასთან.უნდა აღინიშნოს, რომ ამ შემთხვევაშიც სტატიის ავტორებმა სიფრთხილე გამოიჩინეს დაამ შედეგით დაკმაყოფილდნენ, ანუ თავსებადობა აღნიშნეს, მაგრამ ჰიპოთეზისსტატისტიკური შემოწმება აღარ დაუწყიათ.
სხვადასხვაბირთვული გენების ანალიზის საფუძველზე აგებული „ხეები“ (მიტოქონდრიული დნმ–ის„ხის“ მსგავსი) მიანიშნებენ, რომ საერთო წინაპარი (ხის „ფესვები“) 100 ათას წელზეუფრო დიდი ხნის წინ არსებობდა. სხვაგვარად რომ ვთქვათ, „ფესვების“ ასაკი უფრომეტია, ვიდრე „ჩანაცვლების“ ხანა. მაგალითად, აღმოჩენილია ჰემოგლობინის გენები,რომელთაც აშკარად „აზიური წარმოშობა“ აქვთ, მათი ასაკი კი 200 ათას წელსაღემატება. არის ისეთი გენიც, რომელიც ასევე „აზიური წარმოშობისაა“, მაგრამ მისი„ფესვების“ ასაკი 2 მილიონ წელს უახლოვდება. ამრიგად, არსებობს მონაცემები,რომლებიც „ჩანაცვლების“ ჰიპოთეზით (ყოველ შემთხვევაში, მისი თანამედროვეფორმულირებით) უბრალოდ ვერ აიხსნება.
ცნობილიასტატისტიკური მეთოდები, რომელთა მეშვეობითაც შეიძლება ექსპერიმენტული მონაცემების შეფასება მათი თავსებადობის თვალსაზრისით ორალტერნატიულ მოდელთან. მაგალითად, ჩანაცვლების მოდელის თანახმად, 100 ათას წელზეუფრო „ხნიერი“ გენის „ფესვების“ აფრიკული წარმოშობის ალბათობა 1–ია (ანუ 100%). მულტირეგიონალურიმოდელის მიხედვით კი, თუმცა აფრიკული წარმოშობა ყველაზე უფრო სავარაუდოა, ასეთიალბათობა მხოლოდ 0.5–ია (ანუ 50%). სამწუხაროდ, ამ შემთხვევაში ვერც მიტოქონდრიულდნმ–ისა და ვერც Y-ქრომოსომის ანალიზი დამაჯერებელ პასუხს ვერ იძლევა,რადგან ორივე მათგანი მხოლოდ „ერთი გენეტიკური ერთეულია“ (წრიული მიტოქონდრიული დნმჟავის ერთი მოლეკულა, ერთი წრფივი Y-ქრომოსომა), დასტატისტიკისათვის სანდო მასალას ვერ მოგვცემს.
ადამიანისწარმომავლობის მოდელების განხილვის გაგრძელებასა და ამ მიმართულებით კვლევისგანვითარების ანალიზს შემდეგ სტატიაში შემოგთავაზებთ.
ილუსტრაციები:
ნახ. 1: ბირთვული(მარცხნივ) და მიტოქონდრიული (მარჯვნივ) დნმ–ის მემკვიდრეობით გადაცემისმექანიზმების შედარება.
ნახ. 2: ადამიანისევოლუციის „მულტირეგიონალური“ (მარცხნივ) და „აფრიკიდან ჩანაცვლების“ (მარჯვნივ)მოდელები.
ნახ. 3: ადამიანებისგანსახლების შესაძლო სქემა „აფრიკიდან ჩანაცვლების“ მოდელის თანახმად.
ნახ. 4: ადამიანისწარმოშობის „მულტირეგიონალური“ მოდელი.
სტატიის წყარო: გაზეთი 24 საათი
მიტოქონდრიული დნმ:
მოლეკულური ანალიზი და დასკვნები ევოლუციის შესახებ
წინა სტატიაში მიტოქონდრიული დნმ–ის ანალიზისსაფუძველზე წარმოქმნილი მოდელების განხილვა დავიწყეთ. გავიხსენოთ, რომმიტოქონდრიულ დნმ–ს, ბირთვულისაგან განსხვავებით, ინდივიდები მხოლოდ დედისაგანიღებენ. შესაბამისად, მიტოქონდრიული დნმ ერთი თაობიდან მეორეში მდედრობითი სქესისსაშუალებით გადაეცემა – დედისაგან ყველა შვილზე, მაგრამ შემდეგ თაობას(შვილიშვილებს) ამ დნმ–ს მხოლოდ ქალიშვილები გადასცემენ. მიტოქონდრიული დნმ–ისანალიზის საფუძველზე შესაძლებელია ადამიანთა დიდი ჯგუფების წარმომავლობის დადგენა,რადგან მიტოქონდრიული დნმ–ის საერთო მახასიათებლები და მისი განსხვავება სხვაჯგუფებისაგან მდედრობითი სქესის საერთო წინაპრის (მეტაფორულად – „ევას“) არსებობასმიანიშნებენ. ასეთი ანალიზის საფუძველზე წარმოიქმნა „აფრიკიდან ჩანაცვლების“ („Out-of–Africa replacement“) მოდელი, რომლის თანახმადაც თანამედროვე ადამიანის წინაპრებიაფრიკიდან გამოვიდნენ და სხვა კონტინენტებზე არსებული ჯგუფების ჩანაცვლებისშედეგად სათავე დაუდეს ყველა თანამედროვე ადამიანს. რამდენიმე უზუსტობის მიუხედავად,რომელთა განხილვა წინა სტატიაში დავიწყეთ და ახლა გავაგრძელებთ, ეს თეორიაგაცილებით უკეთ მიესადაგება არსებულ მონაცემებს, ვიდრე ე.წ. „მულტიცენტრული“მოდელი, რომლის თანახმადაც ადამიანის ჯგუფებს განსხვავებული წინაპრები ჰყავთ,რომელთა ევოლუცია პარალელურად ხდებოდა (მაგ., სხვადასხვა კონტინენტებზე).
„გენეტიკურიმანძილები“ და ეკოლოგიური შეცდომები.რამდენიმე სტატიაში, რომელიც 2000–იანი წლების შუაში გამოქვეყნდა, გენეტიკურიმანძილები (ანუ ერთი და იმავე წარმოშობის გენებს შორის არსებული სხვაობა) ამჯერადბირთვული გენების მაგალითზე იყო გამოთვლილი. ერთ–ერთ სტატიაში აღწერილი გამოთვლებისსაფუძველზე, ავტორები ასკვნიდნენ, რომ ბირთვული გენების უმეტესობა (~80%) ავლენდა„არააფრიკული წარმოშობის“ გენებთან ურთიერთქმედების ნიშნებს. ეს არ ნიშნავს, რომჩვენი გენების 80% არააფრიკული წარმოშობისაა, მაგრამ დასკვნის მიხედვით, აფრიკიდანგამოსული წინაპრების გენებმა მნიშვნელოვანი ცვლილება განიცადა ადგილობრივარააფრიკულ პოპულაციებთან (ჯგუფებთან) ურთიერთობის შედეგად. ამ ნაშრომის სათაურშიმკაფიოდ იყო ნათქვამი: „გენომური კვლევები უარყოფს, რომ ადამიანის წარმოშობამხოლოდ აფრიკიდან მოხდა“.
მეორე სტატიაში კი, რომელიც იმავე წელს (2005)რამდენიმე დღის დაგვიანებით დაიბეჭდა, ავტორები ამტკიცებდნენ, რომ მათ მიერჩატარებული კომპიუტერული ანალიზი სრულიად თავსებადი იყო „აფრიკიდან ჩანაცვლების“მოდელთან. მათი მტკიცებით, აქ სრული ჩანაცვლება მოხდა, ანუ „არააფრიკული გენების“მოქმედების კვალი თანამედროვე ადამიანში უბრალოდ ნოლს უტოლდებოდა.
რასაკვირველია, მეტად საინტერესოა, როგორ შეიძლებაამ ურთიერთგამომრიცხავი შედეგების ახსნა? ორივე სტატიას ხომ გულმოდგინედ დაგეგმილიდა გულდასმით შესრულებული კომპიუტერული გამოთვლები ედო საფუძვლად. ამ შემთხვევაშიცრამდენიმე აშკარა ნაკლულოვნება გვხვდება: რაც მთავარია, არცერთ სტატიაში არაამოცემული ალტერნატიული ჰიპოთეზის შემოწმება, ანუ არაა ნათქვამი, შეიძლება თუ არაიგივე შედეგები საწინააღმდეგო ჰიპოთეზით აიხსნას, და რადმენად დამაჯერებელი იქნებაეს ახსნა სტატისტიკური თვალსაზრისით. ერთ–ერთ სტატიაში უბრალოდ ნათქვამია, რომ„ჩანაცვლების“ მოდელის გამოყენებით ეს შედეგები ოთხჯერ უკეთ აიხსნება, ვიდრემულტირეგიონალური მოდელ(ებ)ის დახმარებით. მაგრამ ის არაა აღნიშნული, რომ„ჩანაცვლების“ მოდელით მიღებული შედეგების მხოლოდ 10% აიხსნება (შესაბამისად,მულტირეგიონალური მოდელ(ებ)ით – 2.5%). ეს შეფარდება შეიძლება მიუთითებდეს, რომჩანაცვლების მოდელი სხვებს სჯობია, მაგრამ მისი „უნივერსალურობის“ საბუთად ვერ გამოდგება.ასევე, უპასუხოდ დარჩა ერთი – სტატისტიკურად მეტად მნიშვნელოვანი – კითხვა:საკმარისია თუ არა ეს ოთხმაგი განსხვავება, რომ რამდენიმე ალტერნატიული მოდელისშედარების „დამაჯერებლობა“ 5%–ს აღემატებოდეს? 5% ნებისმიერი ბიოლოგიურიექსპერიმენტის ზღვრულ ალბათობად ითვლება, ანუ თუ რაიმე შედეგის შემთხვევით მიღებისალბათობა 5%–ს აღემატება, მას სანდო მონაცემად ვერ მივიჩნევთ. ამ სტატიებში ესმნიშვნელოვანი კითხვა უპასუხოდაა დარჩენილი. ამგვარად, ორივე სტატია შემოიფარგლებაავტორების რჩეული მოდელის შესაბამისობის დამტკიცებით ამათუიმ ჰიპოთეზასთან.
ასევე მნიშვნელოვანია, თუ რატომ იყო მიღებული ამ ორსტატიაში ასე დიამეტრალურად განსხვავებული მონაცემები – და რატომ გახდაშესაძლებელი მიუკერძოებული ანალიზის შედეგად ორი სხვადასხვა მოდელის მისადაგება.ერთ–ერთი შესაძლო ახსნა იმაშია, რომ ამ სტატიების ავტორებს გამოთვლების დასაწყისშირამდენიმე განსხვავებული დაშვება აქვთ გაკეთებული. ასეთი დაშვებების მაგალითებია:ადამიანის რამდენი პოპულაციაა აღებული ანალიზისათვის, რა სიჩქარით მიმდინარეობსმათი ზრდა, ხდება თუ არა ამ ზრდის დროს გენების დრეიფი(ანუ მათი გავრცელებისსიხშირის მკვეთრი ცვლილება) და ა.შ. სავსებით შესაძლებელია, რომ ორ სტატიაში ესპარამეტრები მნიშვნელოვნად განსხვავდება. ასევე, ერთი ნაშრომის მოდელი ასახავდაგადარჩევას ისეთი ნიშან–თვისებების სასარგებლოს, რომლებიც თანამედროვე ადამიანსახასიათებს, მეორის მოდელირება კი ეყრდნობოდა ნეიტრალურ ევოლუციას (ანუ ისეთ პროცესს, როდესაც ხდება მუტაციებისდაგროვება, მაგრამ ჯერ არაა გარკვეული, სასარგებლო თუ საზიანოა ისინისახეობისათვის).
ეს შემთხვევა დამაჯერებლად გვიჩვენებს მოდელირებისსუსტ მხარეს: განსხვავებული დაშვებების გამო შეიძლება გამოთვლის შედეგებიცმკვეთრად განსხვავდებოდნენ, განსაკუთრებით მაშინ, როდესაც ეს დაშვებები უცნობ(ზოგჯერ კი პრინციპულად განუსაზღვრელ) პარამეტრებს ეხება. უპასუხოდ რჩება კითხვა –ამგვარ სტატიებში ორი განსხვავებული მოდელის შედარება ხდება თუ მხოლოდ და მხოლოდსხვადასხვაგვარი დაშვებების გავლენის შესწავლა გამოთვლების შედეგებზე? სანამ ამკითხვაზე დამაჯერებელი პასუხი მოიძებნება, გაურკვეველია, შეიძლება თუ არაგენეტიკური მანძილებისა და მრავალფეროვნების მონაცემების გამოყენება ადამიანის ევოლუციისთაობაზე არსებული ჰიპოთეზების შესამოწმებლად.
არსებობს კიდევ ერთი ფაქტორიც – ე.წ. „ეკოლოგიურიშეცდომა“, რომელიც ყველა ნაშრომშია დაშვებული და რომელიც ხდება, როდესაც მკვლევარებსმიაჩნიათ, რომ პროცესების ერთობლიობის მოდელის სტატისტიკური დამტკიცება საკმარისიაამ პროცესების საფუძველში მდებარე ან მათი გამომწვევი მექანიზმების მოდელისჭეშმარიტების დასამტკიცებლად. ასეთი დამოკიდებულება რამდენიმე მიზეზის გამოამცდარი: 1) არ არსებობს პირდაპირი და ერთმნიშვნელოვანი დამოკიდებულება საწყისპროცესებსა და მათ საფუძველზე მომხდარ შედეგებს შორის; 2) შეუძლებელია უკლებლივ ყველასაწყისი პროცესის განსაზღვრა, რომლის განვითარებაც ერთი და იმავე შედეგითაა დაგვირგვინებული; 3) საწყისიპროცესების მოდელირება ძალიან ხშირად მრავალ დაშვებაზეა დამოკიდებული; 4)გამოთვლები მხოლოდ პროცესის გვიანდელ ეტაპებს ეხება და არა საწყის საფეხურებს, რისშედეგადაც შესაძლებელია უნებლიე „მიკერძოება ერთობლიობის მიმართ“ (aggregationbias).
ამ მოზასრებების საფუძველზე, ნებისმიერი, თუნდაცუზადოდ შესრულებული მოდელირებით დამაჯერებლად ვერ შემოწმდება, მიმდინარეობდა თუარა ადამიანის ევოლუციის პროცესის საწყისი ეტაპები რომელიმე კონკრეტული მექანიზმისსაფუძველზე. მოდელირება სავსებით გამოსადეგია შესაძლებელი მექანიზმებისგამოსარკვევად ანდა სხვა მოდელირებული პროცესების უარსაყოფად, მაგრამ მოდელისჭეშმარიტების დადგენა ამ გზით უბრალოდ შეუძლებელია. მაგალითად, ზოგიერთშემთხვევაში მოდელირების საფუძველზე შეიძლება დავასკვნათ, რომ გარკვეულიმონაცემები არ მიესადაგება „აფრიკიდან ჩანაცვლების“ ჰიპოთეზას, მაგრამ ამდებულებიდან მულტირეგიონალური ჰიპოთეზის ჭეშმარიტება არ გამომდინარეობს, თუნდაც იმუბრალო მიზეზის გამო, რომ ეს უკანასკნელი ერთადერთ შესაძლო ალტერნატივას არწარმოადგენს.

როგორც ამ მოდელებიდან და მათი განხილვიდან ჩანს,ადამიანის წარმოშობისა თუ განსახლების, და ამ მოვლენების საფუძველში მდებარემექანიზმების თაობაზე აზრთა სხვადასხვაობა არსებობს. თანამედროვე ადამიანისწარმოშობისა თუ განსახლების თარიღებზე (უფრო ზუსტად, ამ თარიღების „თანრიგზე“) კიმეცნიერული (ანუ ფაქტებით შემოწმებული და დადასტურებული) მონაცემები საკმაოდმკაფიოდაა დადგენილი. ნათელია, რომ ეს მრავალი ათეული ათასი წლის წინ მოხდა:საწყისი გამოსვლა აფრიკიდან – დაახლოებით 1.8 მილიონი წლის წინ, ხოლო „აფრიკიდანჩანაცვლება“ – 100 ათას წელზე უფრო ადრე (150–200 ათასი წლის წინ). არსებობს დამატებითი მოსაზრებაც – რომ მეორე გამოსვლააფრიკიდან 65 ათასი წლის წინ დაიწყო. ისიცცხადია, რომ მითოლოგიური თუ ლიტერატურული ცნობები იმის თაობაზე, რომ ადამიანი 6ათასი წლის წინ წარმოიშვა (თუ შეიქმნა), უბრალო მეტაფორად ან ავტორ(ებ)ისეულიწარმოსახვის ნაყოფად უნდა აღვიქვათ.
და მაინც, რატომ მოხდა ადამიანთა განსახლებააფრიკიდან დაახლოებით 60 ათასი წლის წინ, ანუ მათი წარმოშობიდან 100–150 ათასიწლით უფრო გვიან? ერთ–ერთ მოდელს, რომელიც ამის ახსნას ცდილობს, აქ განვიხილავთ.როგორც უკვე აღვნიშნეთ, გენეტიკურად და ანატომიურად თანამედროვე ადამიანის სახეობაუკვე არსებობდა აფრიკაში სულ ცოტა 150 ათასი წლის წინ. ასევე დადგენილია, რომ სულცოტა 40 ათასი წლის წინ დასრულდა ამ სახეობის განსახლება აფრიკიდან და მის მიერსხვა, „არქაული“ ჯგუფების ჩანაცვლება (მაგ., ნეანდერტალელებისა, რომლებიც იმდროისათვის ევროპაში ცხოვრობდნენ). ამ ზოგადი ცნობების მიუხედავად, რამდენიმეძირეული კითხვა იმის შესახებ, თუ რატომ და როგორ მოხდა ეს განსახლება დაჩანაცვლება, საკმაოდ დიდხანს უპასუხოდ რჩებოდა.
ამ მიმართულებით კვლევისას ორი მნიშვნელოვანიკითხვა არსებობს:
1) ცნობილია, რომ გენეტიკურადდა ანატომიურად თანამედროვე ადამიანთა პოპულაციები (ინდივიდთა ერთობლიობები,რომლებიც სხვა ამგვარი ერთობლიობებისაგან მეტ–ნაკლებად გამოყოფილნი არიან) უკვეარსებობდნენ აფრიკაში სულ ცოტა 150 ათასი წლის წინ. მაშინ რატომ გავიდა კიდევ ~100ათასი წელი, სანამ ეს პოპულაციები აფრიკიდან გამოვიდნენ და მსოფლიოს სხვარეგიონებში (ანუ სხვა კონტინენტებზე) განსახლდნენ?
2) რა ევოლუციურიპროცესის გავლენით „მოახერხეს“ ამ პოპულაციებმა სრულიად ახალი და უცხოტერიტორიების ათვისება? რაც არანაკლებ საინტერესოა, როგორ მოხდა, რომ ამტერიტორიებზე მრავალი წლის განმავლობაში მცხოვრები „არქაული“ პოპულაციები, რომლებიცადგილობრივ პირობებთან უკვე შეგუებულნი იყვნენ, დამარცხდნენ „ახალმოსულებთან“ბრძოლაში?
როგორცაღმოჩნდა, შესაძლო პასუხი ამ კითხვებზე მოინახა თანამედროვე აფრიკული პოპულაციებისდნმ–ის ანალიზის საფუძველზე, ისევე როგორცსამხრეთ და აღმოსავლეთ აფრიკაში აღმოჩენილი არქეოლოგიური კვლევების შედეგად.
აფრიკული დნმ–ის მონაცემები. დემოგრაფიულირეკონსტრუქციის მიმართულებით ჩატარებული კვლევები მეტად მრავალფეროვან და ზოგჯერურთიერთსაწინააღმდეგო შედეგებს იძლევა. ეს დებულება ყველა ადამიანის ყველათანამედროვე პოპულაციას ეხება, და მონაცემები აფრიკის პოპულაციის შესახებგამონაკლისს არ წარმოადგენს. დემოგრაფიული ისტორიის ნაკვალევთა ანალიზი რთულდებამწვავე დისკუსიებით მრავალ თემაზე: მუტაციების წარმოქმნის არაერთგვაროვანი ტემპისხვადასხვა გენებში, სელექციის გავლენა დნმ–ის მოლეკულურ შემადგენლობაზე,მიგრაციებით გამოწვეული გართულებები დემოგრაფიულ სტრუქტურაში, და ასე შემდეგ.როგორც უკვე აღვნიშნეთ, მიტოქონდრიული დნმ–ის კვლევის საფუძველზე მიღებულიშედეგებს რამდენიმე აშკარა უპირატესობა აქვთ: ამ დნმ–ის მემკვიდრეობით გადაცემამხოლოდ დედის მხრიდან ხდება, მუტაციების ანალიზი გაცილებით უფრო ინფორმატიულშედეგებს იძლევა ბირთვულ დნმ–თან შედარებით, გარემოს გავლენით გადარჩევის გავლენამცირეა. სწორედ მიტოქონდრიული დნმ–ის ანალიზის შედეგად გაირკვა, რომ ჩვენსწინაპართა აფრიკული პოპულაციები მკვეთრად გაიზარდა 80 ათასიდან 60 ათასამდე წლისწინ, ანუ დაახლოებით 100 ათასი წლის დაგვიანებით იმის შემდეგ, რაც თანამედროვეაფრიკული პოპულაციების უგვიანესი საერთოწინაპარი (most recent common ancestor, MRCA) წარმოიშვა.
ამ ჰიპოთეზისმხარდამჭერი შედეგი პირველად მიიღეს მიტოქონდრიული დნმ-ის შემადგენლობაში არსებულიგანსხვავებების ანალიზით (mismatch analysis). იგულისხმება განსხვავებები,რომლებიც ერთი და იგივე პოპულაციის ფარგლებში არჩეული ორი ინდივიდის შორისარსებობს. ამ განსხვავებების სიხშირის პოპულაციებში გავრცელების შესწავლის შედეგადგამოირკვა, რომ აფრიკულ პოპულაციებში ყველაზე ხშირი ისეთი განსხვავებები იყო,რომელთა წარმოშობის „ასაკი“ დაახლოებით 80 ათას წელს შეადგენდა. აზიური დაევროპული პოპულაციებში კი უდიდესი სიხშირით, შესაბამისად, 60 და 40 ათასი წლის წინწარმოქმნილი განსხვავებები აღმოჩნდა. რასაკვირველია, ამ განსხვავებების „ასაკი“მხოლოდ მიახლოებითია და გამოითვლება იმ ვარაუდის საფუძველზე, თუ რა სიხშირითწარმოიქმნება მიტოქონდრიულ დნმ-ში მუტაციები (დნმ-ის შემადგენელი ერთეულებისცვლილებები). მაგრამ რახან ეს ვარაუდი სამივე პოპულაციის ანალიზს თანაბრად მიესადაგება, ამშედეგების ერთობლიობიდან გამომდინარეობს, რომ: 1) აფრიკული პოპულაციებისრაოდენობის სწრაფი ზრდა გაცილებით უფრო ადრე მოხდა, ვიდრე აზიასა და ევროპაში. ესდასკვნა იმ მონაცემების საფუძველზე კეთდება, რომ „აფრიკული მუტაციების მრავალფეროვნება“ უფრო „ხნიერია“, ვიდრე სხვაკონტინენტებზე. 2) აფრიკის მოსახლეობის ზრდის შედეგად მოხდა მოსახლეობის„ტალღისებრი გავრცელება“ საწყისი, შედარებით მცირე რეგიონიდან, ამავე კონტინენტისსხვა ნაწილებზე.
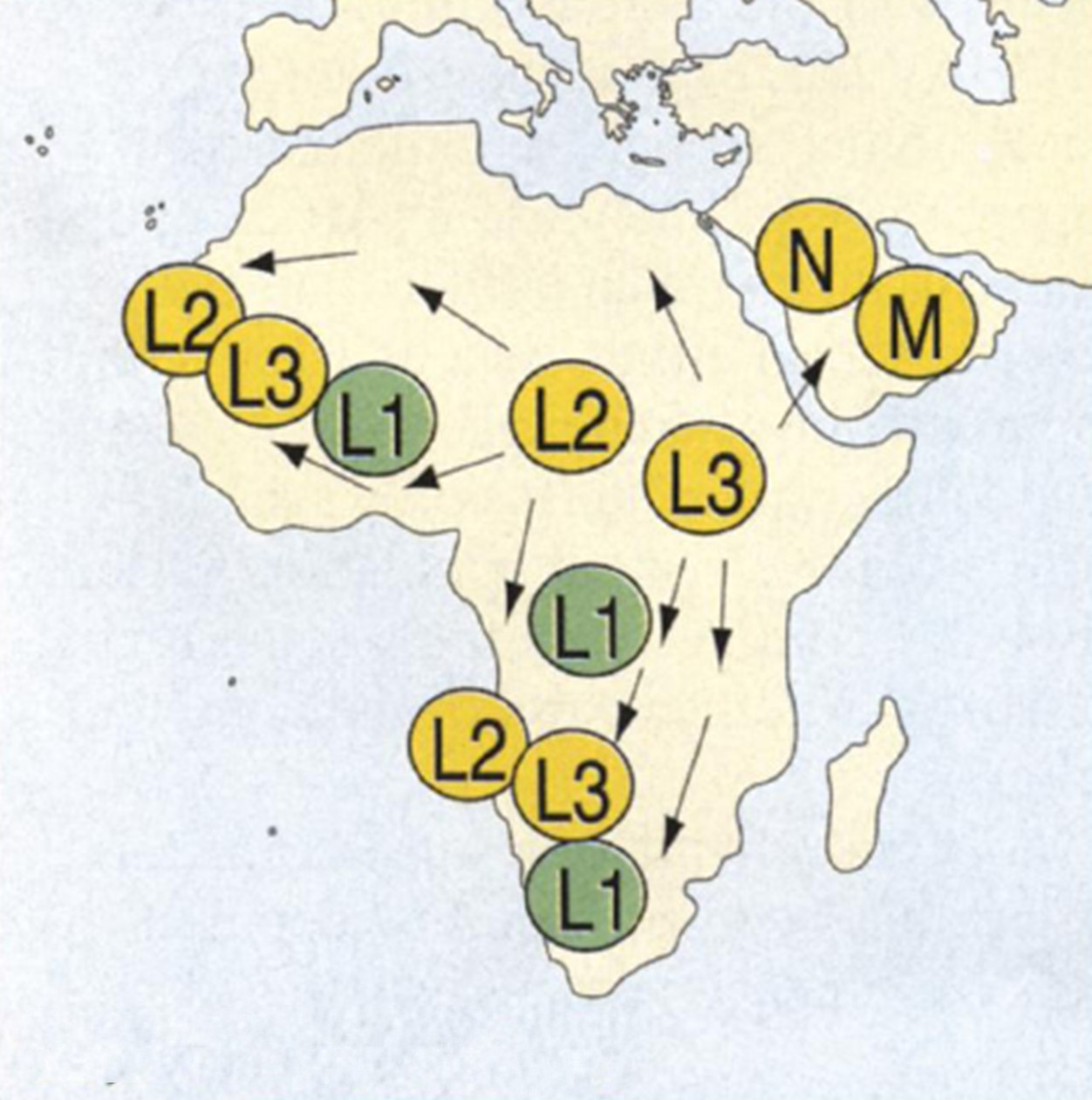
რამდენიმე წლის წინ ამ ვარაუდის მორიგი მხარდამჭერი მონაცემები დაგროვდა თანამედროვეაფრიკული პოპულაციების მიტოქონდრიული დნმ-ის ანალიზის შედეგად. გარკვეულიშემადგენლობის (L2 და L3) მიტოქონდრიული დნმ-ის მნიშვნელოვანი გავრცელება 80ათასიდან 60 ათასი წლის წინანდელი დროითაა განსაზღვრული. ამ შედეგების თანახმადაც,განსახლება დაიწყო აღმოსავლეთი ან სამხრეთი აფრიკის შედარებით მცირე რეგიონიდან.შემდგომ, მოსახლეობის გადინება დასავლეთი აფრიკის მიმართულებითაც დაიწყო და ამრეგიონს 30-40 ათასი წლის წინ მიაღწია. აზიის იმ რეგიონებში კი, რომლებიც აფრიკისსაზღვარზეა განლაგებული, განსახლება 60-65 ათასი წლის წინ მოხდა (შესაძლოა, წითელიზღვის შემოვლის გზით). ნებისმიერ შემთხვევაში, როგორც მიტოქონდრიულ დნმ-ში არსებული განსხვავებებისანალიზი, ასევე L2 და L3 შემადგენლობის გავრცელების შესწავლა მიანიშნებს, რომადამიანთა განსახლება დაიწყო შედარებით მცირე გეოგრაფიული რეგიონიდან, და ესგანსახლება პოპულაციის რაოდენობის მკვეთრი ზრდის შედეგად მოხდა.
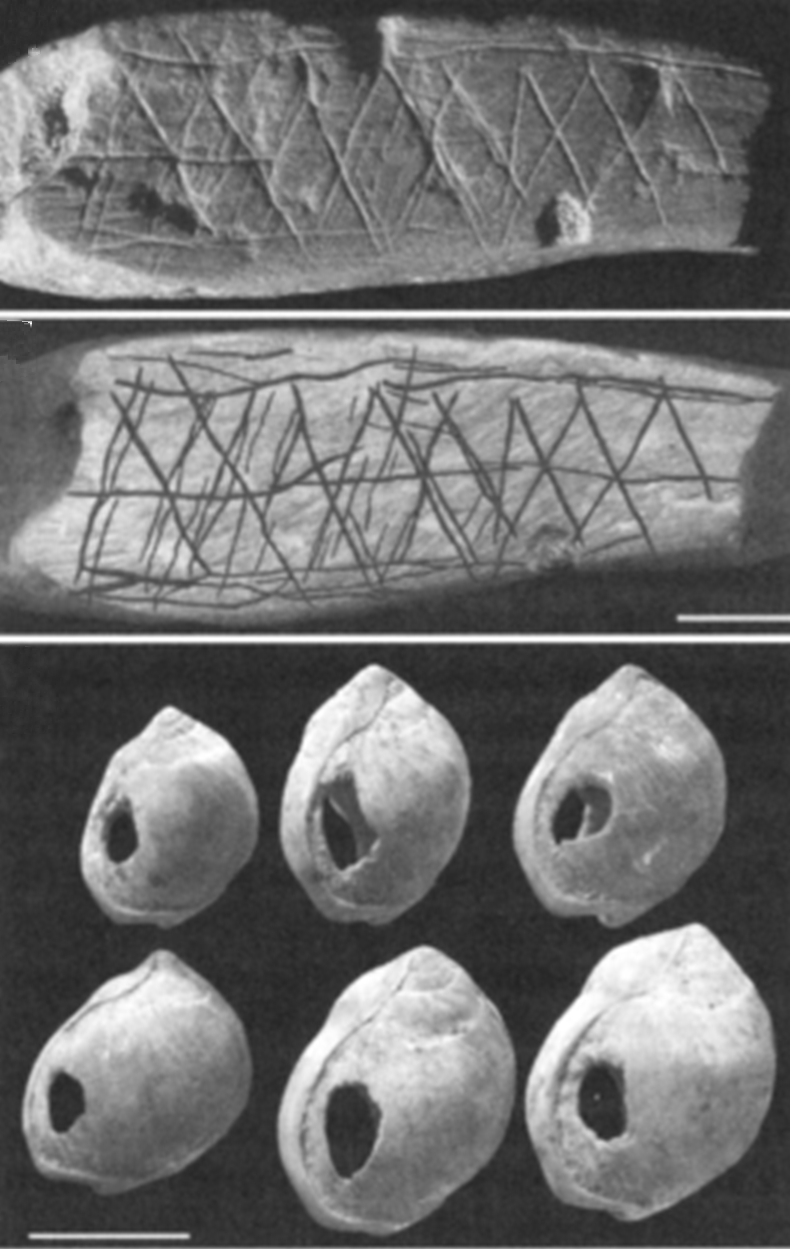
არქეოლოგიური კვლევის შედეგები. რამ გამოიწვია აფრიკულიპოპულაციების რაოდენობის ასეთი მკვეთრი ზრდა 60-80 ათასი წლის წინ? მოსალოდნელია,რომ სამხრეთ და ცენტრალურ აფრიკაში ჩატარებული არქეოლოგიური გათხრები ამ კითხვაზეპასუხს გაგვცემენ. ამ რეგიონებში აღმოჩენილი საგნები ძირითადად შუა ქვის ხანასგანეკუთვნებიან (250 ათასიდან 40 ათას წლამდე). ამასთანავე, ზოგიერთ მათგანში უკვეაშკარად ჩანს რადიკალური ტექნოლოგიური და კულტურული წინსვლის ნიშნები, რომლებითაცისინი მკვეთრად გამოირჩევიან შუა ქვის ხანის ტიპური ფორმებისაგან და რომელნიც უფროწააგავენ ევროპასა და აზიაში ნაპოვნ ნივთებს, რომლების ასაკი 45-50 ათასი წლითგანისაზღვრება (ე.წ. „ზედა პალეოლითის რევოლუციის“ პერიოდი). ასეთია, მაგალითად,ნაჯახის გამოკვეთის განსხვავებული ტექნოლოგია, ტყავის, ხისა და ძვლისდამუშავებისათვის განკუთვნილი ხელსაწყოები, ძვლისა ან ქვისაგან დამზადებულიიარაღები და სხვა. ამავე პერიოდს მიეკუთვნება ხელოვნების პირველი ნიმუშებიც -ორნამენტები ძვლებსა და ნიჟარებზე. ყველა ეს ნივთი იმის მიმანიშნებელია, რომ ამდროს ხდებოდა ტექნოლოგიის სწრაფი განვითარება, რაც უმნიშვნელოვანეს სიახლესწარმოადგენდა შუა ქვის ხანის პერიოდისათვის.
ილუსტრაციები:
ნახ. 1:L2 და L3 მიტოქონდრიულიდნმ-ის გავრცელების სავარაუდო მიმართულებები 80 ათასი – 60 ათასი წლის წინ.
ნახ. 2:იარაღები და ხელსაწყოები,რომლებიც 65 ათასი წლის წინაა დამზადებული და რომლებიც არქეოლოგებმა სამხრეთაფრიკაში აღმოაჩინეს. ასეთივე ხელსაწყოები, რომლებიც ევროპასა და აზიაშიააღმოჩენილი, გაცილებით უფრო გვიანდელია – ისინი 45 ათასი წლით თარიღდება, რაც„აფრიკიდან განსახლების“ პროცესის მიმანიშნებელია.
ნახ. 3:75 ათასი წლის წინანდელიხელოვნების ნიმუშები, რომლებიც სამხრეთ აფრიკაშია აღმოჩენილი.
სტატიის წყარო: გაზეთი 24 საათი
ადამიანის გენების მოლეკულური ანალიზი:
რა მოხდა 500, 100, ან 60 ათასი წლის წინ, და როგორ ვიგებთ ამისშესახებ?
წინა სტატიებში მიტოქონდრიული დნმ–ის ანალიზის საფუძველზეწარმოქმნილი მოდელები და ადამიანის აფრიკიდან განსახლების სხვადასხვა დროის დამიმართულებების მხარდამჭერი საბუთები განვიხილეთ. წინამდებარე (ამ თემისათვისდასკვნითი) სტატია ადამიანის წარმოშობისა და განვითარების შესწავლისას მიღებულახალ შედეგებს დაეთმობა.
გავიხსენოთ, რომ მიტოქონდრიები ადამიანისუჯრედსშიდა წარმონაქმნებია (ორგანელები), რომლებიც ბირთვის გარეთ მდებარეობს დარომელთა შიგთავსში არსებული დნმ (გენეტიკური მასალის განმსაზღვრელი მოლეკულა)მომავალ თაობას მხოლოდ დედის მხრიდან გადაეცემა. ამის გამო მისი ცვალებადობაგაცილებით ნაკლებია, ვიდრე ბირთვული დნმ–ისა, რომელსაც ყველა ადამიანი როგორცდედისგან, ისე მამისაგან იღებს. ამავე კანონზომიერების საფუძველზე, ადამიანთა დიდიჯგუფების წარმომავლობის ანალიზი გაცილებით იოლია მიტოქონდრიული დნმ–ისგამოყენებით, რადგან საერთო წინაპრის (მეტაფორულად „ევას“) არსებობის შემთხვევაშიმათი მიტოქონდრიული დნმ–ის შემადგენლობა ერთმანეთთან გაცილებით უფრო ახლოა, ვიდრეგანსხვავებული წარმოშობის ჯგუფის წევრებისა.
ასეთი კვლევების შედეგად წარმოქმნილი მოდელისთანახმად, ანატომიურად თანამედროვე ადამიანის წარმოქმნა აფრიკაში მოხდა, ~200ათასი წლის წინ. აფრიკიდან განსახლება კი გაცილებით უფრო გვიანი პროცესია – მხოლოდ65 ათასი წლის წინანდელი.
გასულ ზაფხულს კი ადამიანის წარმომავლობისშესწავლის დარგში კიდევ ერთი საინტერესო აღმოჩენა გაკეთდა: გამოირკვა, რომაფრიკული და არააფრიკული პოპულაციების (ინდივიდთა ერთობლიობის) შორის საკმაოდდიდხანს ხდებოდა გენეტიკური მასალის მიმოცვლა – 60 ათასი წლის წინ მომხდარი„აფრიკიდან ჩანაცვლების“ (Out-of-Africareplacement) შემდეგაც კი. ეს კი ნიშნავს, რომშეჯვარება ამ ჯდუფებს შორის აფრიკიდან საწყისი გამოსვლის შემდეგ კიდევ დიდხანსმიმდინარეობდა.
ამ კვლევაში პირველად მოხდა ცალკეული გენომების(ადამიანთა გენების ერთობლიობის) შესწავლა. წინამორბედ კვლევებში კი არა მთელიგენომის ანალიზი, არამედ რამდენიმე გენის შესწავლა ხდებოდა. ამიტომ საჭირო იყო ამგენების ანალიზი მრავალ ადამიანში, რომდასკვნები უფრო დამაჯერებელი ყოფილიყო. ახალმა მეთოდმა კი საშუალება მისცა არქეოლოგებს, ინდივიდების გენების საფუძველზე გაეკეთებინათდასკვნები მათ წინაპართა პოპულაციების სიდიდეზე და განვითარების ისტორიაზე. ეს კი,თავის მხრივ, იძლევა შესაძლებლობას, ნაკლები რაოდენობით დაშვება თუ შეუმოწმებელივარაუდი გაკეთდეს ამ პროცესების მოდელირებისას. ასევე, ამ მეთოდით უკეთგამოირკვევა ადამიანის წარმომავლობა და მისი, როგორც ბიოლოგიური სახეობის,განვითარების ისტორია 1 მილიონიდან 10 ათასამდე წლის წინ.
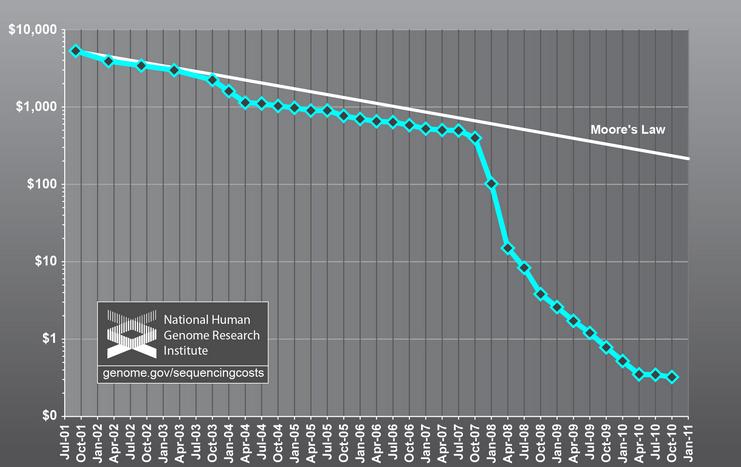
რასაკვირველია, თვით ასეთიანალიზის შესაძლებლობა განაპირობა დნმ-ის შემადგენლობის ანალიზის (სეკვენირების)ტექნოლოგიის მნიშვნელოვანმა განვითარებამ, გამარტივებამ და, რაც მთავარია,გაიაფებამ.უკანასკნელი დებულების საილუსტრაციოდ ერთ მაგალითს მოვიშველიებთ: სულ რაღაც10 წლის წინ სეკვენირების სიძვირე (~10ათასი აშშ დოლარი დნმ-ის 1 მილიონი ერთეულის – ნუკლეოტიდის – თანამიმდევრობისგასარკვევად) მასობრივი ანალიზის ჩატარებას მნიშვნელოვან დაბრკოლებას უქმნიდა.ახლა კი იგივე ოპერაციის ჩატარების ღირებულება 0.1 დოლარს უახლოვდება. სხვაგვარადრომ ვთქვათ, თანამედროვე ტექნოლოგია საშუალებას გვაძლევს, 1 აშშ დოლარად 10მილიონი ნუკლეოტიდის შემცველი დნმ ფრაგმენტის სეკვენირება შევასრულოთ.
ხსენებულ ნაშრომში 7 ინდივიდის მთლიანიგენომის ანალიზი ჩატარდა: 3 ევროპელი, ჩინელი, კორეელი და 2 დასავლეთ აფრიკელიმამაკაცის გენომი იყო სეკვენირებული. აღმოჩნდა, რომ თუმცა აფრიკული და არააფრიკულიპოპულაციების ერთმანეთისაგან გამოყოფა 120 – 100 ათასი წლის წინ დაიწყო, ისინიფაქტობრივად ერთ პოპულაციად რჩებოდნენ ჯერ კიდევ 80-60 ათასი წლის წინ.
ამის შემდეგ ევროპასა და აღმოსავლეთაზიაში მოხდა ადამიანის წინაპართა პოპულაციების რიცხვის მნიშვნელოვანი შემცირება -საწყის რაოდენობასთან შედარებით 10%-მდე. ეს მოვლენა კიდევ ერთ მნიშვნელოვანპერიოდს ემთხვევა: ევროპისა და აზიის პირველი თანამედროვე ადამიანების ნამარხიფორმებისა და არტეფაქტების ასაკი სწორედ ამ დროით განისაზღვრება. ამასთანავე, ისიც გამოირკვა, რომ სულ ცოტა 20ათასი წლის განმავლობაში „აფრიკიდან გამოსულსა“ და „აფრიკაში დარჩენილ“პოპულაციებს შორის ჯერ კიდევ მიმდინარეობდა შეჯვარება და, მაშასადამე, გენებისმომოცვლა. ამ პროცესის შესაძლო ახსნა იმაშია, რომ აფრიკიდან მოსული „ახალი ემიგრანტების“შერევა „აფრიკიდან ჩანაცვლების“ (წინა ტალღის) წარმომადგენლებთან საკმაოდ დიდიხნის განმავლობაში ხდებოდა.
ამრიგად, ამ კვლევის საფუძველზე რამდენიმესაყურადღებო დასკვნა გაკეთდა: 1) ადამიანის პოპულაციების რაოდენობამ მნიშვნელოვნადიმატა აფრიკაში დაახლოებით 100 ათასი წლის წინ, როდესაც იქ თანამედროვე ადამიანიგაჩნდა; 2) ევროპასა და აზიაში მცხოვრები არააფრიკული წარმოშობის ინდივიდების შესწავლამ აჩვენა,რომ 60 ათასი წლის წინ მათი წინაპრების პოპულაციები მნიშვნელოვნად შემცირდა. ესშედეგი მოულოდნელი არ იყო, რადგან მსგავსი დასკვნა ადრინდელი კვლევებისსაფუძველზეც გაკეთდა. ახალი შედეგი კი ისაა, რომ 3) ამის შემდეგ დაახლოებით 20ათასი წლის განმავლობაში მიმდინარეობდა გენების მიმოცვლა (ანუ შეჯვარება)ადგილობრივსა და აფრიკიდან გამოსულ ინდივიდებს შორის.
თანამედროვე ადამიანებისადა მათი არცთუ შორეული ნათესავების (მაგ., ნეანდერტალელების) პოპულაციებისურთიერთქმედებაც ევოლუციური ანალიზის არანაკლებ საინტერესო საკითხია. ამ პრობლემისკვლევის ერთ–ერთი მეთოდია X-ქრომოსომის ანალიზი. გასული წლის ზაფხულშიგამოქვეყნებული კიდევ ერთი ნაშრომის შედეგების საფუძველზე, ზოგიერთი ადამიანის X-ქრომოსომისნაწილი ნეანდერტალელების X-ქრომოსომისაგან წარმოსდგება და, რაც განსაკუთრებითსაყურადღებოა, ამ ადამიანთაგან უკლებლივ ყველა აფრიკის ფარგლებს გარეთ ცხოვრობს…
ნეანდერტალელებისწინაპრებმა აფრიკა 800 ათასიდან 400 ათასამდე წლის წინ დატოვეს. მათი ევოლუციათანამედროვე ევროპაში გრძელდებოდა, ახლანდელისაფრანგეთის, ესპანეთის, გერმანიისა და რუსეთის ტერიტორიაზე. ნეანდერტალელები ამადგილებზე ჯერ კიდევ 30 ათასი წლის წინ ცხოვრობდნენ. ამასთანავე, თანამედროვე ადამიანთაპირველი წარმომადგენლები აფრიკიდან 80 ათასიდან 50 ათასამდე წლის წინ გამოვიდნენ. ნეანდერტალელებიადამიანებზე ფიზიკურად უფრო ძლიერნი იყვნენ, ამასთანავე ლაპარაკის უნარი გააჩნდათდა, როგორც არქეოლოგები ვარაუდობენ, ფლეიტისმაგვარ მუსიკალურ ინსტრუმენტზე დაკვრაცკი შეეძლოთ. ამ ორი სახეობის კვლევისას წარმოიქმნა ერთი მეტად საინტერესოპრობლემა: მართლაც წარმოადგენდნენ თუ არა ნეანდერტალელები ცალკეულ სახეობას თუხდებოდა მათი შეჯვარება ადამიანებთან? უკვე ცნობილია, რომ ამ კითხვაზე დადებითიპასუხი არსებობს: ადამიანებსა და ნეანდერტალელებს შორის ახლო ურთიერთობაარსებობდა.
დაახლოებით ათი წლის წინმონრეალის უნივერსიტეტის (University of Montreal, Canada) მკვლევართა ჯგუფმააღმოაჩინა ფრაგმენტი ადამიანის X-ქრომოსომაში, რომელიც ამ ქრომოსომის სხვაშენადგენელი ნაწილებისაგან გამოირჩეოდა და რომლის წარმოშობამაც მეცნიერთაგანსაკუთრებული ინტერესი გამოიწვია. 2010 წელს ამ საკითხის კვლევაში მნიშვნელოვანწარმატებას მიაღწიეს: ნეანდერტალელების გენომის სრული სიკვენსი (ანუ დნმ–ისშემადგენელი ერთეულების თანამიმდევრობა) განისაზღვრა. ამ შედეგის მიღებისთანავე,მონრეალელმა მეცნიერებმა დაადგინეს, რომ ადამიანის X-ქრომოსომის უჩვეულო ფრაგმენტინეანდერტალელების X-ქრომოსომის ნაწილს შეესაბამებოდა. მეტიც, ამ ფრაგმენტის 6000სხვადასხვა ადამიანის X-ქრომოსომებთან შედარებამ კიდევ უფრო შთამბეჭდავი შედეგიმოიტანა: „ნეანდერტალელების თანამიმდევრობა“ (the Neanderthal sequence) არსებობდაყველა კონტინენტის მცხოვრებთა დნმ–ში, ავსტრალიის ჩათვლით. გამონაკლისს მხოლოს ქვე–საჰარულიაფრიკის (sub-Saharan Africa) მცხოვრებლები შეადგენდნენ.
ამ შესანიშნავი შედეგისსაფუძველზე თითქმის უეჭველია, რომ ამ ფრაგმენტის არსებობისა და გავრცელების მიზეზიიმაშია, რომ ჩვენი (თანამედროვე ადამიანების) წინაპრების და ნეანდერტალელებისშორის შეჯვარება ხდებოდა. მონაცემების დამაჯერებლობას ის გარემოებაც განამტკიცებს,რომ კვლევის დასაწყისისას ცნობილი სულაც არ იყო, რომ უჩვეულო ფრაგმენტი„ნეანდერტალული“ წარმოშობისა იყო: ეს მხოლოდ მოგვიანებით გამოირკვა,ნეანდერტალელების გენომის სრული ანალიზის შემდეგ! სხვაგვარად რომ ვთქვათ, ავტორებმა ჯერ საინტერესო ფრაგმენტის არსებობა და„განსაკუთრებულობა“ დაადგინეს, დასკვნები მისი სავარაუდო წარმოშობის შესახებ კიმოგვიანებით, დამოუკიდებელი კვლევის შედეგებთან შედარების საფუძველზე გაკეთდა.
როგორც ადვილიწარმოსადგენია, ამ სტატიების გამოქვეყნებიდან სულ რამდენიმე თვის შემდეგ გაჩნდასარეკლამო განცხადება, რომელიც მკითხველს სთავაზობდა მოლეკულური ანალიზისსაფუძველზე გაერკვია მასში „ნეანდერტალელის გენების“ წილი. როგორც განცხადებისავტორები ამტკიცებენ, ადამიანთა უმეტესობა ნეანდერტალელების დნმ–ის 2.5% შეიცავს.„გამოწვევა“, რომელიც კლიენტებს ეგზავნებათ, მდგომარეობს იმის გარკვევაში, 2.5%–ზემეტი თუ ნაკლები „ნეანდერტალური გენეტიკური ინფორმაცია“ აქვს ამათუიმ პიროვნებას.რასაკვირველია, მეტად ძნელი წარმოსადგენია, რომ ამ ანალიზის შედეგებთან რაიმეგარეგნული ნიშან–თვისებების დაკავშირება შეიძლება (მაგალითად, გონებრივიშესაძლებლობების, სხეულზე არსებული თმის საფარის, და ა.შ.), თუმცა სავარაუდოა, რომამგვარი მასიური ანალიზის ჩატარება რომელიმე საინტერესო კანონზომიერებასგამოავლენს ადამიანის ევოლუციის „მოლეკულურ მატიანეში“.

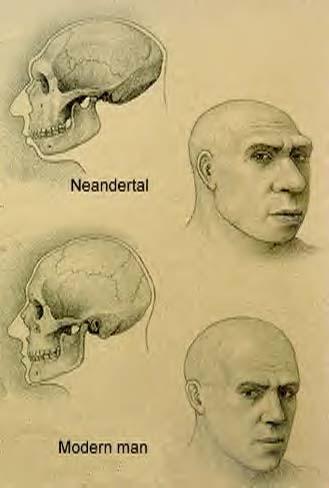
გარეგნული ნიშან–თვისებებისსხვაობის შესახებ ადამიანთა და ნეანდერტალელთა შორის არსებობდა მოსაზრება, რომთავის ქალის სხვადასხვა ფორმა სხვადასხვა გარემო პირობების გავლენით (და,მაშასადამე, ბუნებრივი გადარჩევის წნეხით) არის გაპირობებული: ნეანდერტალელთამოგრძო სახე აიხსნება მათ მიერ კბილების, როგორც იარაღის, გამოყენებით;თანამედროვე ადამიანთა „შეწეული“ (retracted) სახე კი ამ აუცილებლობისგაუჩინარების, ისევე როგორც მეტყველების განვითარების შედეგია. როგორც დევისისუნივერსიტეტის (University of Davis, California, USA) არქეოლოგებმა დაადგინეს,სახის ფორმის განსხვავებები გაცილებით უკეთ აიხსნება გენებში შემთხვევითიცვლილებების დაგროვებით იმ >500 ათასი წლის განმავლობაში, როდესაცნეანდერტალელები და თანამედროვე ადამიანების წინაპრები ერთმანეთისაგან იყვნენიზოლირებულნი. გავიხსენოთ, რომ ნეანდერტალელები აფრიკიდან 800 ათასიდან 400ათასამდე წლის წინ გამოვიდნენ, თანამედროვე ადამიანები კი მხოლოდ 65 ათასი წლისწინ, და ამ ორი – დამოუკიდებლად ჩამოყალიბებული – პოპულაციის ურთიერთქმედებამხოლოდ ამ დროიდან დაიწყო.
გენეტიკური ანალიზი დასტატისტიკური მეთოდები აგრეთვე გამოიყენება თანამედროვე ადამიანთა ცალკეუკიჯგუფების ანალიზისათვის. კორნელის უნივერსიტეტის (Cornell University, USA) მეცნიერებმა6 ადამიანის სრული გენომის ანალიზი ჩაატარეს. ამ კვლევაში შესწავლილი იყოაღმოსავლეთ აზიის, ევროპის, დასავლეთი და სამხრეთი აფრიკის წარმომავლობისადამიანთა გენები. ამ ნაშრომის ძირითადი დასკვნის თანახმად, სანის (San)სამხრეთაფრიკული ტომი, რომელიც ნადირობითა და შემგროვებლობით ირჩენს თავს,ადამიანის სხვა პოპულაციებს გაცილებით უფრო ადრე გამოეყო, ვიდრე აქამდე იყომიჩნეული. ახალი კვლევის თანახმად, ეს 130 ათასი წლის წინ მოხდა. შედარებისათვისგავიხსენოთ, რომ თანამედროვე ევროპული პოპულაციების წინაპრები აფრიკიდან მხოლოდ 60ათასი წლის წინ გამოვიდნენ.
ამ ნაშრომშიც მთავარი როლიადამიანის გენომის სრულ ანალიზს ენიჭება. ადრეული კვლევები, რომლებიცმიტოქონდრიული დნმ–ის ან Y-ქრომოსომის მემკვიდრეობით გადაცემას ეფუძნება, გენებისმხოლოდ მცირე ნაწილს შეისწავლის. ამ შემთხვევებში მემკვიდრეობის მხოლოდ ერთი ხაზიგანიხილება: „ევას“ (მიტოქონდრიული დნმ–ის გადაცემა მხოლოდ მდედრობითი სქესისმიერ, ორივე სქესის შვილებში) ან „ადამის“ (Y-ქრომოსომის გადაცემა – მხოლოდმამრობითი სქესის წარმომადგენლებში). სრული გენომის ანალიზი კი არა ერთი, არამედმრავალი მემკვიდრეობითი ხაზის შესწავლის საშუალებას იძლევა.
დაბოლოს, მოკლედგანვიხილოთ „ნეანდერტალელების გენომის“ პროექტის (NeanderthalGenome Project)არსი, რადგან ეს კვლევა ტექნოლოგიური მიღწევების გამოყენებისა და მიღებულიშედეგების გულმოდგინე ანალიზის შერწყმის შესანიშნავი მაგალითია. გენომის სრულიანალიზი მეტად რთული ამოცანაა თანამედროვე ორგანიზმების შემთხვევაშიც, აქ კი დნმ–ისწყარო 38–44 ათასი წლის ძვლები გახლდათ. ამოცანას ისიც ართულებდა, რომ ამძვლებიდან დნმ–ის გამოყოფის მცდელობისას დიდი რაოდენობით გროვდებოდა იმბაქტერიების დნ მჟავის მოლეკულები, რომლებიც ამ ძვლებზე იმყოფებოდნენ. თვითნეანდერტალელების დნ მჟავა კი მცირე ზომის ფრაგმენტებად იყო დანაწევრებული.ანალიზისათვის შესაფერი დნმ–ის მიღების პროცესისათვის არანაკლებ დაბრკოლებასწარმოადგენდა ამ მოლეკულების ქიმიური მოდიფიკაცია, რაც ასევე ხშირი აღმოჩნდანეანდერტალელების დნმ–ის ანალიზისას.
ასევე საჭირო იყო ნეანდერტალელებისდნმ–ის გამოყოფის სპეციალური მეთოდების შემუშავება, და მისი დაცვა ადამიანისდნმ–ის მინარევებისაგან. სეკვენირების ახალი ტექნოლოგიით შესაძლებელი გახდაგამოყოფილი დნმ–ის პირდაპირი ანალიზი – მისი გამრავლების გარეშე, რამაც შესაძლოშეცდომებისა თუ არტეფაქტების რიცხვი მნიშვნელოვნად შეამცირა. მეორე მხრივ, ამმეთოდის გამოყენებით მიღებული შედეგების გადამოწმება (ანუ მრავალჯერადისეკვენირება) არც ისე იოლია, მაგრამ მიღებული მონაცემები საკმარისი აღმოჩნდა, რომარსებულ გენომებთან (ადამიანისა და შიმპანზეს) ინფორმატიული შედარებითი ანალიზიჩატარებულიყო. მაგალითად, ზოგიერთ ფრაგმენტებში, რომელნიც ადამიანისა და შიმპანზესშორის განსხვავდება, ნეანდერტალელებს ჯერ კიდევ შენარჩუნებული აქვთ მსგავსებაშიმპანზესთან… ამის საფუძველზე, შესაძლებელი გახდა ისეთი გენეტიკური ცვლილებებისგამოაშკარავება და კლასიფიკაცია, რომლებიც თანამედროვე ადამიანის წარმოშობას დაედოსაფუძვლად.
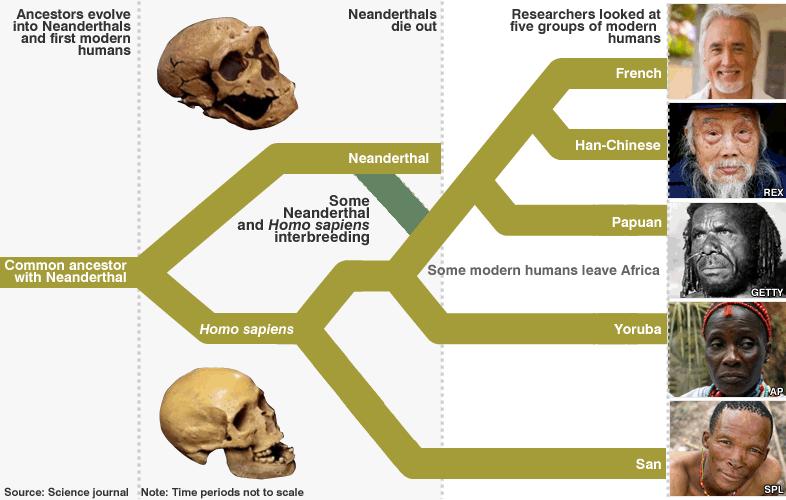
ვარაუდობენ, რომ ადამიანისადა შიმპანზეს წინაპართა სახეობები ერთმანეთისაგან განცალკევდნენ 5–6 მილიონი წლისწინ. ნეანდერტალელებისა და ადამიანების წინაპრები, აღწერილ შედეგებზე დაყრდნობით,440 ათასიდან 270 ათასამდე წლის წინ „გაიყვნენ“.
ადამიანებსა დანეანდერტალელებს შორის გენების შედარებით გვიანდელი მიმოცვლა იმითაც მტკიცდება,რომ თანამედროვე ადამიანების გენომის მსგავსება ნეანდერტალელებთან ინდივიდუალურია(იხ. ზემოთ). ეს მიმოცვლა რომ სახეობების გაყოფისთანავე შეწყვეტილიყო, მაშინ ყველათანამედროვე ადამიანის განსხვავება ნეანდერტალელისაგან ერთი და იგივე იქნებოდა. ესკი სინამდვილეს არ შეეფერება – როგორც უკვე აღვნიშნეთ, არააფრიკული პოპულაციებისმსგავსება ნეანდერტალელებთან გაცილებით მეტია, ვიდრე აფრიკული მოსახლეობისა დანეანდერტალელების შედარების საფუძველზე მიღებული.
არსებული განსხვავებებისმიზეზი ამ ორ სახეობებს შორის არსებულ შეჯვარებათა მცირე რაოდენობაც კი შეიძლებაგამხდარიყო. მეცნიერთა გამოთვლების თანახმად, გენების მიმოცვლა 80 ათასიდან 50 ათასამდეწლის წინ ხდებოდა. ეს პროცესი, სავარაუდოდ, მაშინ დაიწყო, როცა ადამიანთაწინაპრები აფრიკიდან განსახლდნენ და ნეანდერტალელებს პირველად შეეყარნენ.
რაც შეეხება ამპოპულაციების შორის არსებულ კულტურულ კავშირებს, ამის შესახებ დასაბუთებულივარაუდის გაკეთება უბრალოდ შეუძლებელია. მთავარი აღმოჩენა, რომელიც ამ კვლევებისშედეგად გაკეთდა, გენების მიმოცვლის მხარდამჭერი საბუთების გამოაშკარავებაშიმდგომარეობს.
დასკვნა: დნმ–ის თანამიმდევრობის(სიკვენსის) ანალიზის შესაძლებლობამ და ტექნოლოგიურმა მიღწევებმა ამ სფეროში ადამიანისწარმომავლობის ისტორიას მრავალი მნიშვნელოვანი დეტალი დაუმატა. მეთოდები, რომლებიცსულ რაღაც ორიოდე ათეული წლის წინ წარმოუდგენლად გვესახებოდნენ, ახლა თითქმისრუტინულ პროცედურებად იქცნენ. რაც მთავარია, ჩვენი შორეული წინაპრებისწარმომავლობასა თუ მათი განსახლების გზების შესახებ გაკეთებული დასკვნები სულ უფრონაკლებად ემყარება დაშვებებსა თუ ვარაუდებს და სულ უფრო მეტად – გენებისა დაქრომოსომების ანალიზის შედეგად მიღებულ ექსპერიმენტულ შედეგებს. ეს კი, თავისმხრივ, ჩვენი წარმომავლობისა თუ განვითარების ისტორიის ცალკეულ დეტალებზე უფროზუსტი კითხვების დასმისა და მათზე უფრო დასაბუთებული პასუხების გაცემისშესაძლებლობას გვაძლევს.
ილუსტრაციები:
ნახ. 1: დნმ–ის სეკვენირების ღირებულების შემცირების ამსახავი სქემა. მურის კანონისთანახმად, ღირებულება წრფივად უნდა მცირდებოდეს, მაგრამ ამ შემთხვევაშისეკვენირების ფასი გაცილებით უფრო მკვეთრად იკლებს.
ნახ. 2, 3: ნეანდერტალელისა და თანამედროვე ადამიანის თავის ქალების შედარება და ამშედარების საფუძველზე აღდგენილი სახის ნაკვთები.
ნახ. 4: თანამედროვე ადამიანების წარმოშობისა და განვითარების სქემანეანდერტალელების როლის გათვალისწინებით.
სტატიის წყარო: გაზეთი 24 საათი

")
")





